Abrus precatorius - a deadly beauty
Gerhard Prenner, researcher in plant morphology and anatomy, presents his recent studies on Abrus precatorius, a "deadly beauty" with fascinating flowers and inflorescences.


The genus Abrus consists of around 17 species distributed across Africa, Madagascar, India and Indo-China. Among them, the jequirity bean (Abrus precatorius L.) is probably the best and widest known species. It is a weed found throughout the tropics and neotropics.
Some of its many common names, such as bead vine, black-eyed Susan or coral bead plant point to the use of its hard, shiny black and red seeds for stringing into necklaces and rosaries in India and other tropical countries. This is also documented in Kew's Economic Botany collection, which houses several necklaces and other decorative items with seeds of Abrus.

However, the seeds are also well known because they contain the highly poisonous protein abrin. This is a plant lectin, related to ricin, that inhibits cellular protein synthesis. Therefore Abrus precatorius might be called a "deadly beauty" and wearing bracelets and other adornments made from jequirity beans is not recommended.
Systematists have placed the genus Abrus in its own tribe Abreae, which is found in a relatively isolated systematic position closely related to another tribe of legumes called Millettieae (Lewis et al., 2005). To contribute to a better understanding of this taxon, I performed detailed analyses of its inflorescence (Prenner, 2013a) and flowers (Prenner, 2013b) using Kew's scanning electron microscope (SEM), plant material from Kew's herbarium collections and literature from Kew's extensive library collections. These studies provide new and exciting insights not only into the evolution and systematic placement of this fascinating taxon but also in a wider context among the big and economically important bean family.
Legume research at Kew
Kew has a long history of research into the bean family, or Leguminosae. With more than 19,500 species in 751 genera, this is not only one of the largest plant families on earth (ranked third after orchids and the daisy family) but it is also one of the most important plant groups, providing many protein-rich foods for our planet's ever increasing population. Beans, lentils, soya, chickpeas, peanuts and peas are just some of the many prominent staple foods produced by the family.
Over the last 40 years, scientists at Kew have played a leading role in legume research, describing many new and potentially important species (and a noteworthy number of new genera), and in providing valuable insights into the systematic relationships, morphology and chemistry of the group. The well-curated herbarium, which houses more than 750,000 legume specimens, provides the baseline material central to this research and is regularly visited by scientists and students from around the world.

Kew also houses an extensive collection of legume material preserved in alcohol, wood samples in the Economic Botany collection, seeds in the Millennium Seed Bank, valuable legume literature and, last but not least, an impressive number of living legume species in the gardens and green-houses. Together these collections underpin legume research at Kew, and have helped me to conduct detailed research on the flowers and inflorescence of Abrus precatorius, the results of which I present below.
Abrus precatorius: Flowers
Flowers of Abrus deviate from the common floral ground plan of the majority of the bean family in that only nine stamens (instead of ten) are found in the mature flower. Therefore one aim was to see what happened to the "lost" stamen. Is it initiated but lost in a later developmental stage or is it completely lost without any detectable primordial evidence? Answering this question is important in order to evaluate this character correctly and to put this uncommon floral feature in a broader context of the family.
A close inspection of early developmental stages showed that, in the position where normally a tenth stamen would be found, no traces of an organ appear. Interestingly, the two filaments of the stamens adjacent to this gap grow inwards and seemingly close the gap.
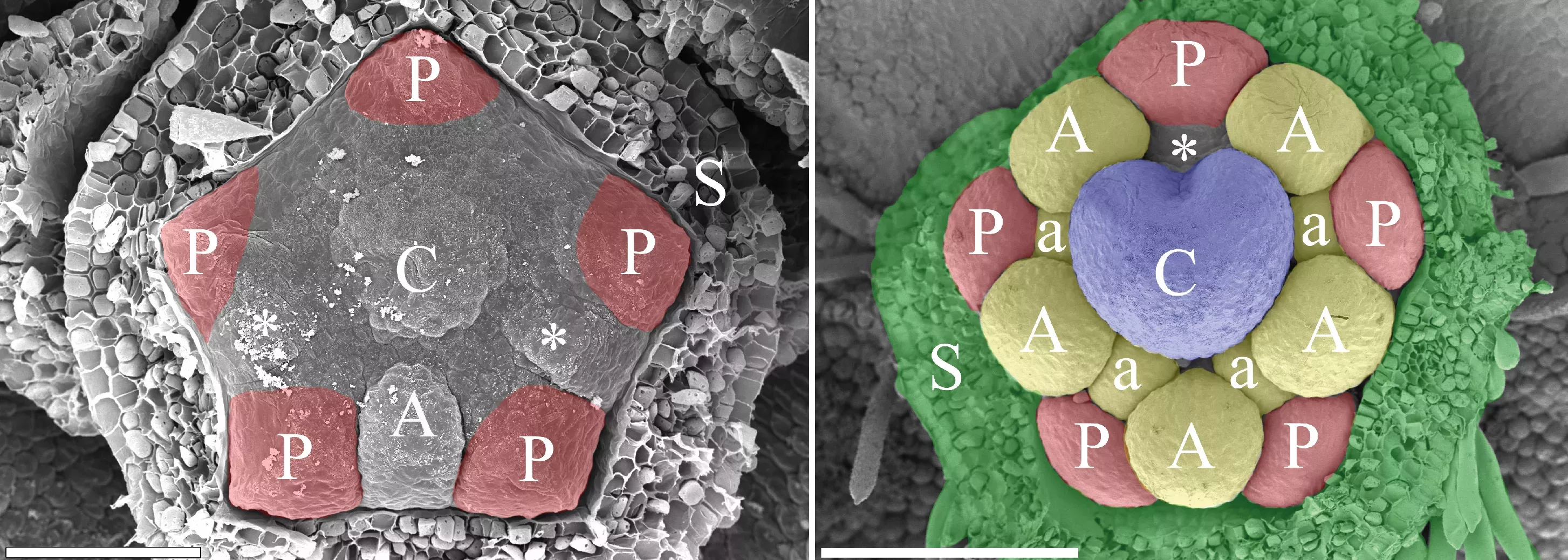
The images above show the following: Left: An early developmental stage showing the simultaneous formation of five petals (red). Right: young flower with all organs formed (calyx removed) showing the gap (asterisk) where a tenth stamen would be formed. Calyx green, petals red, stamens yellow, carpel blue. Scale bar = 0.05 left and 0.1mm right. (Photos Gerhard Prenner)
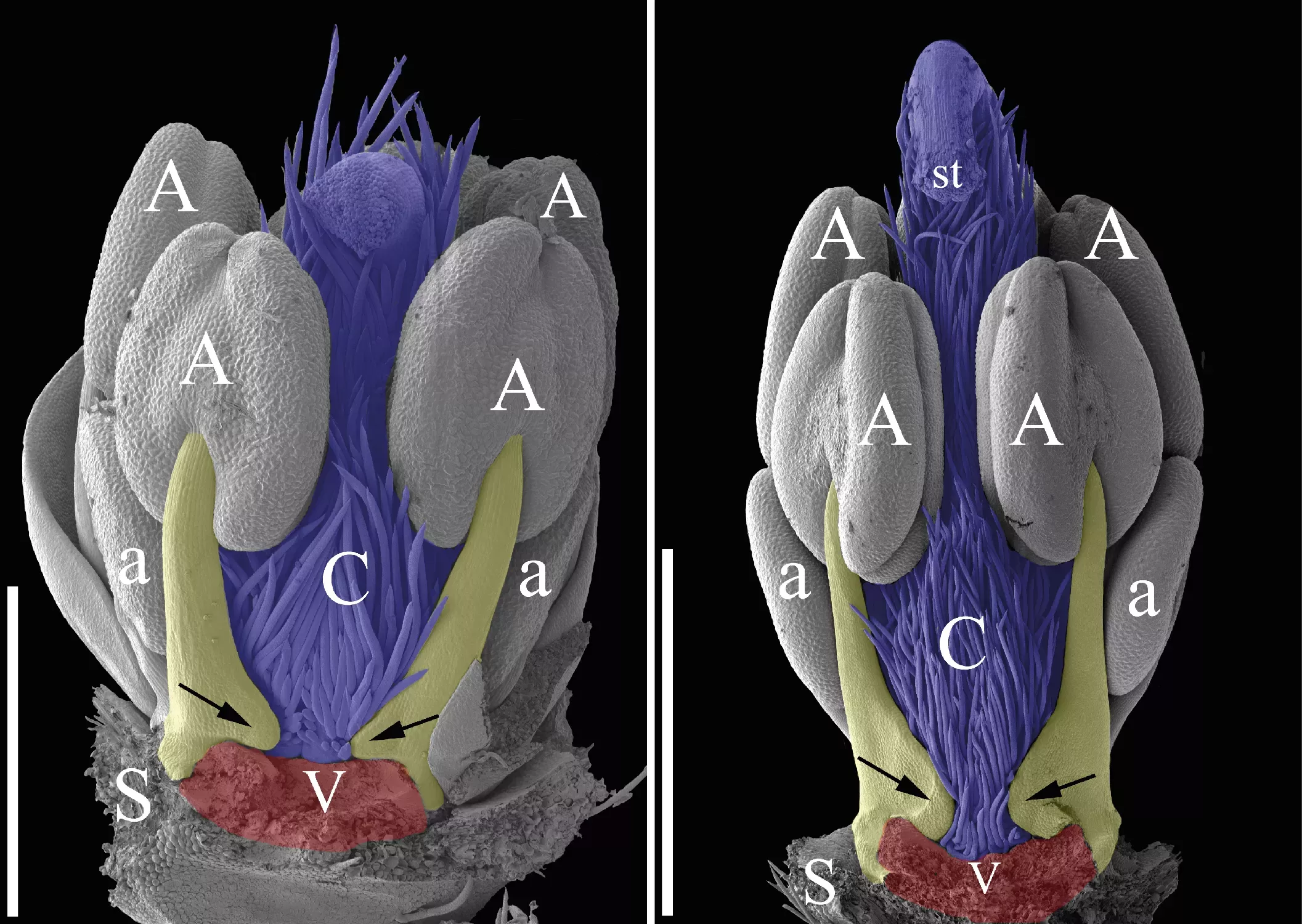
The images above are of two young flowers, showing the gap in front of the petal (red) where normally a stamen would emerge. Instead of this the two stamen filaments of the adjacent stamens (coloured in yellow) are growing inwards towards the gap (arrows). Scale bars = 0.5 mm (left) and 1 mm (right). (Photos: Gerhard Prenner)
One possible explanation for this unusual pattern is that the enlarging filaments provide stability for the flower and compensate for the lost stamen. The remaining nine stamens develop in a normal fashion. Their filaments fuse and form a filament sheath in which the single carpel lies.
Pollen grains are formed within the anthers and are typical of Leguminosae. On the anther surface I found characteristic pores or stomata - a character that is found only occasionally in anthers of the family. The pollen grains show three slits or "colpi" through which pollen will ultimately germinate when it reaches the stigma of another compatible flower.
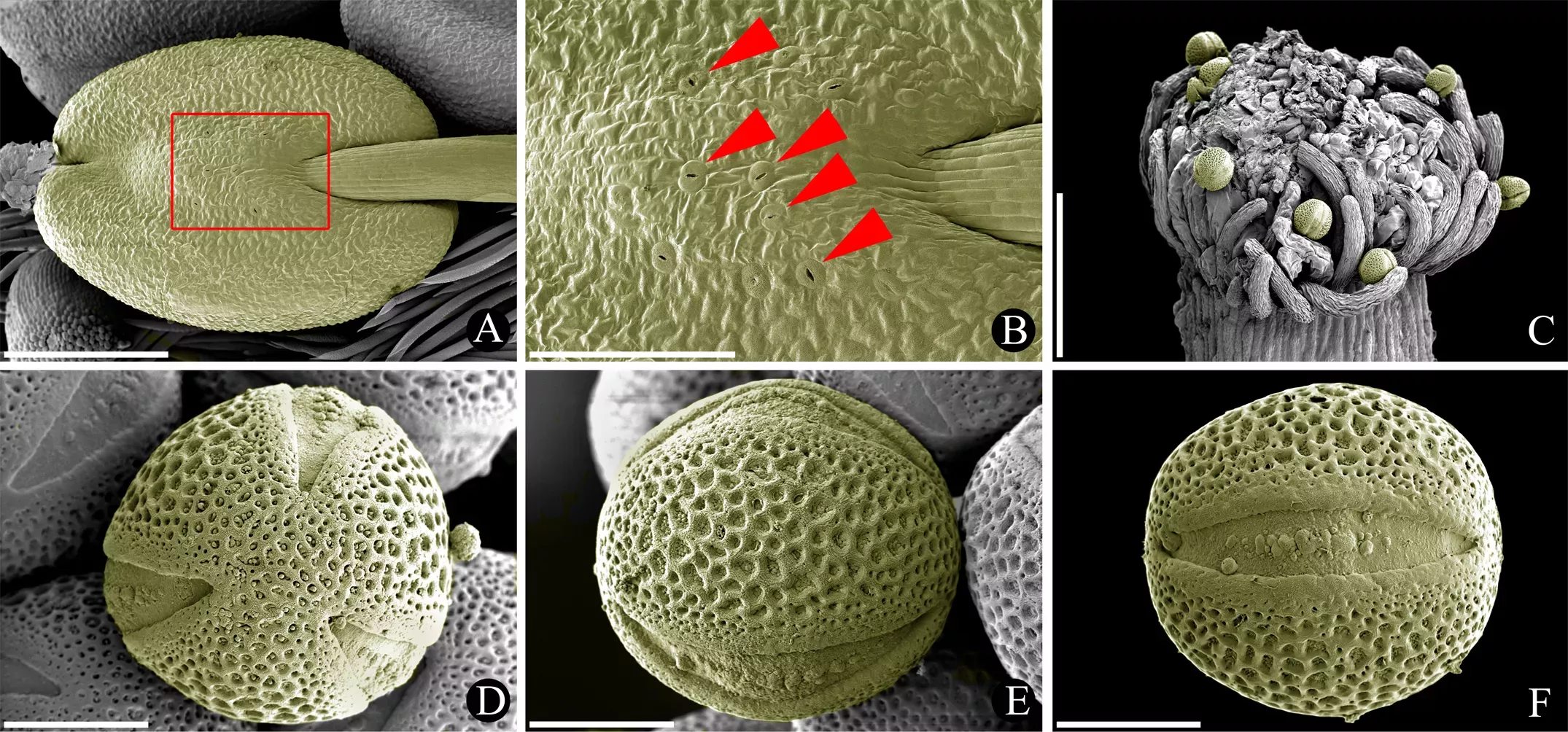
The images above show the following: (A-B) Anther with stomata on its surface. (C) Stigma surrounded by hairs with adhering pollen grains (yellow). (D-E) Pollen grains in three different views: (D) polar view showing the three pores, (E) equatorial view between two pores showing details of the grain's surface and (F) equatorial view on a pore showing details of the pore and the surrounding surface. Scale bar = 0.2 mm in A, 0.1 mm in B, C and 0.010 mm in D-F (Photos: Gerhard Prenner)
Abrus precatorius: Inflorescence
Abrus precatorius has peculiar inflorescences composed of several sub-units (or partial inflorescences) and therefore termed a compound inflorescence. The axis of each partial inflorescence is nodose (with knot-like swellings) and the inflorescence meristem (growth point) is positioned off centre, towards the main inflorescence axis. Individual flowers are formed from the outside towards the centre of this partial inflorescences (= acropetal flower formation) and they arise in a characteristic pendulum pattern switching between right and left. Within each partial inflorescence, a complete series from fully developed flowers to floral primordia can be found (follow the numbers below for the sequence).
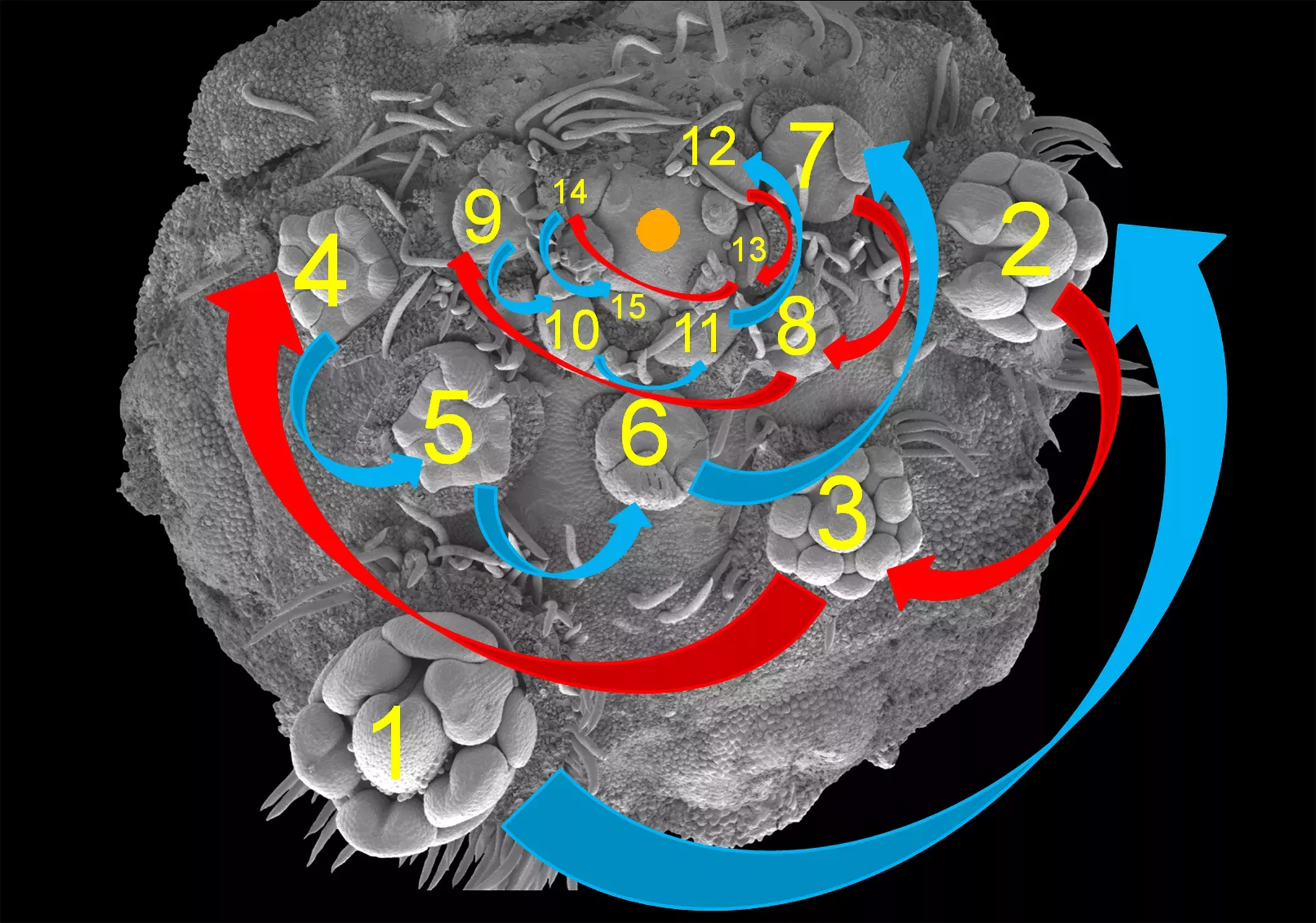
The image above displays a nodose partial inflorescences showing the inflorescence axis on top and off center and also showing the pendulum pattern of flower formation, oscillating between right (blue arrows) and left (red arrows). The axis of the main inflorescences is on top of this figure (marked with orange dot). (Photo: Gerhard Prenner)
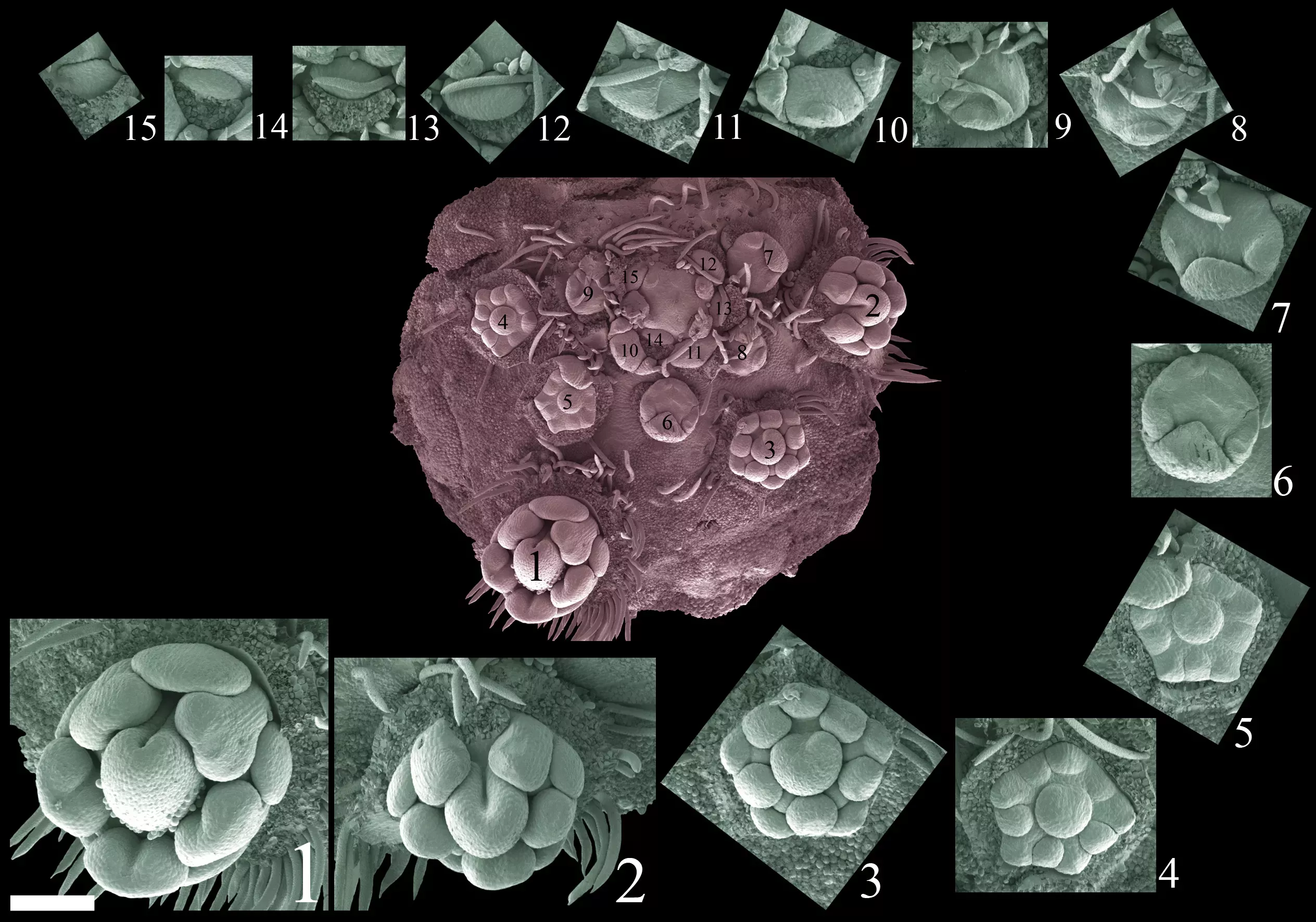
The image above displays a partial inflorescence in red and its individual flowers showing a complete series from perfect flowers (1) to earliest developmental stages (= floral primordia, 15). (Photo: Gerhard Prenner)
These results provide strong evidence that the inflorescence of Abrus can be interpreted as a compound raceme (sensu Prenner et al., 2009) with condensed partial inflorescences. All flowers are formed in a continuous ontogenetic (developmental) series as found in regular racemes of other members of the bean family. The meristem of partial inflorescences was relocated towards the main inflorescence axis. The reason for this relocation might be an inhibitory field acting from the main inflorescence which prevents flower formation close to the main inflorescence axis. The relocation of the meristem might in turn disturb spiral flower formation around a central meristem which is normally found in racemes. Instead of this, flower formation oscillates in a pendulum pattern around the lateral inflorescence axis.
Tribal status
Some ontogenetic patterns such as unidirectional sepal formation and simultaneous petal formation match those found in other species within the tribe Millettieae. In contrast, the peculiar circumstances in the inflorescence and the complete loss of a stamen are rather unusual characters, supporting the isolated position of the genus Abrus and probably justify (along with other characters) its tribal status Abreae.
However, our current knowledge is still only fragmentary. Additional studies will help to clarify and understand not only the systematic placement of the deadly beauty, Abrus precatorius, but also of other important and useful members of the bean family.
References
Lewis, G., Schrire, B., Mackinder, B. & Lock, M. (2005). Legumes of the World. Royal Botanic Gardens, Kew.
Prenner, G., Vergara-Silva, F. & Rudall, P.J. (2009). The key role of morphology in modelling inflorescence architecture. Trends in Plant Science 14: 302–309. Available online
Prenner, G. (2013a). Papilionoid inflorescences revisited (Leguminosae-Papilionoideae). Annals of Botany 112(8): 1567–1576. Available online
Prenner, G. (2013b). Flower development in Abrus precatorius (Leguminosae: Papilionoideae: Abreae) and a review of androecial characters in Papilionoideae. South African Journal of Botany 89: 210–218. Available online


